Cơ chế xác định giới tính các loài
Cho dù một con vật sẽ trở thành một con đực, một con cái hay lưỡng tính đi chăng nữa thì điều đó đã được xác định rất sớm trong quá trình phát triển. Các nhà khoa học đã tìm tòi trong hàng trăm năm để hiểu hệ thống xác định giới tính. Chẳng hạn, vào năm 335 TCN, Aristotle đề xuất rằng sức nóng của người đàn ông trong khi giao hợp giúp xác định giới tính đứa. Nếu sức nóng của nám giới có thể lấn át sự lạnh lẽo của nữ giới, thì một bé trai sẽ được sinh ra và ngược lại. Các giả thuyết về môi trường xác định giới tính, như Aristotle, đã phổ biến cho đến khoảng năm 1900, khi nhiễm sắc thể giới tính được phát hiện. Hóa ra, Aristotle đã tìm ra một thứ gì đó, ít nhất là trong trường hợp của một số loài bò sát, trong đó nhiệt độ của tổ ấp trứng xác định giới tính của phôi. Tuy nhiên, đối với hầu hết các động vật, giới tính được xác định bởi nhiễm sắc thể.
Nhiễm sắc thể giới tính
Xác định giới tính dẫn đến sự phát triển của các cá thể có đặc điểm cho phép họ được xác định là nam, nữ hoặc trong một số trường hợp là lưỡng tính (ở đây tạm nhắc đến loài người làm đại diện). Ở một số loài, như giun tròn C. elegans, sự khác biệt về đặc điểm sinh dục có thể rất nhỏ; trên thực tế, ở C. elegans, đặc điểm phân biệt giới tính duy nhất là sự hiện diện của tinh hoàn hay ống dẫn trứng. Ở các loài khác, sự khác biệt về kiểu hình giữa hai giới có thể khá đáng kể. Ví dụ, hãy xem bộ lông màu mè và bắt mắt của một con gà tây trống so với các đặc điểm khá đơn điệu của một con gà tây mái. Động vật có vú đực và cái cũng dễ dàng được phân biệt bởi nhiều sự khác biệt trong kiểu hình bên trong và bên ngoài, hành vi và sự trao đổi chất của chúng.
Bước đột phá lớn đầu tiên trong việc tìm hiểu xác định giới tính là phát hiện ra nhiễm sắc thể giới tính vào đầu những năm 1900. Từ các phân tích tỉ mỉ trên nhiễm sắc thể côn trùng đực và cái, các nhà khoa học phát hiện ra rằng, mặc dù hầu hết các nhiễm sắc thể có số lượng bằng nhau ở hai giới, có một hoặc hai nhiễm sắc thể xuất hiện không đồng đều giữa hai giới. Các phân tích trên các loài khác trong những năm qua đã tiết lộ rằng sự khác biệt về nhiễm sắc thể chịu trách nhiệm chính cho việc xác định giới tính ở hầu hết các loài động vật.
Cơ chế nhiễm sắc thể của xác định giới tính ở côn trùng
Côn trùng là lớp sinh vật đa dạng nhất trên hành tinh, vì vậy không quá ngạc nhiên khi chúng cho thấy sự đa dạng đáng kể trong cơ chế xác định giới tính của chúng (Saccone et al., 2002). Tuy nhiên, giống như hầu hết các loài động vật khác, phần lớn các loài côn trùng có nhiễm sắc thể giới tính lưỡng hình có thể phân biệt được bằng tế bào học.
Động vật có hai nhiễm sắc thể giới tính khác nhau thuộc giới dị giao tử (heterogametic sex) và do đó chúng có thể tạo ra hai loại giao tử. Ngược lại, các cá thể thể đồng giao tử (homogametic sex) chỉ có thể tạo ra một loại giao tử. Ở người (cũng như nhiều động vật khác), đàn ông thường có một nhiễm sắc thể X và một nhiễm sắc thể Y, trong khi phụ nữ có hai nhiễm sắc thể X. Hệ thống này đảo ngược ở bướm và bướm đêm (Bộ Lepidoptera): con cái thuộc giới dị giới giao tử, trong khi con đực là đồng giao tử. Các nhiễm sắc thể giới tính trong bộ Lepidoptera được ký hiệu là nhiễm sắc thể W và Z. W thường liên quan đến sự phát triển của các đặc tính cái; khi không có nhiễm sắc thể W, ZZ phát triển thành đực và ZO phát triển thành cái. Có nhiễm sắc thể W để phát triển thành cái thậm chí không cần thiết đối với một số loài. Một loài sâu bướm được gọi là Talaeporia tubulosa sử dụng nhiệt độ môi trường để kiểm soát việc xác định giới tính trong trường hợp không có nhiễm sắc thể W (Traut et al., 2007).
Một số châu chấu cũng sử dụng hệ thống xác định giới tính đơn nhiễm sắc thể (XX / XO). Ở đây, con đực chỉ có một nhiễm sắc thể giới tính, vì vậy chúng được coi là XO. Do đó, con đực là giới dị giao tử, vì chúng tạo ra hai loại giao tử khác nhau.
Hệ thống xác định giới tính dựa vào nhiễm sắc thể thậm chí còn tối giản hơn nữa ở một số chi muỗi nhất định, trong đó hai giới … không thể phân biệt được bằng nhiễm sắc thể. Giới tính trong nhóm đồng nhất này được cho là được xác định bởi yếu tố xác định đực trội.
Xác định giới tính ở ruồi giấm
Các nhiễm sắc thể giới tính của ruồi giấm Drosophila melanogaster đã đóng một vai trò đặc biệt quan trọng trong sự hiểu biết của chúng ta về di truyền. Do đó, có thể có một điều gây ngạc nhiên là ruồi giấm sử dụng một cơ chế tương đối hiếm để xác định giới tính. Trên thực tế, ở ruồi giấm, giới tính chủ yếu được xác định theo tỷ lệ X: A hay tỷ lệ số lượng nhiễm sắc thể X với số lượng bộ nhiễm sắc thể (Cline & Meyer, 1996). Sự cân bằng giữa các yếu tố xác định giới cái được mã hóa trên nhiễm sắc thể X và các yếu tố xác định giới đực được mã hóa trên các nhiễm sắc thể thường sẽ quy định mô hình phiên mã đặc trưng giới tính nào sẽ được bắt đầu. Do đó, ruồi XX, XXY và XXYY là con cái, trong khi ruồi XY và XO là con đực. Ruồi không thể tồn tại với hơn hai bản sao của nhiễm sắc thể X vì cơ chế mà chúng sử dụng để bù trừ lượng gen. (Không giống như ở động vật có vú, ở ruồi giấm tất cả các nhiễm sắc thể X vẫn hoạt động và ruồi điều chỉnh lượng sản phẩm của các gen liên kết X bằng cách gấp đôi mức biểu hiện gen trên nhiễm sắc thể X ở con đực. Có thêm một nhiễm sắc thể X, vốn đã chứa gần một phần ba số gen của ruồi, tạo ra tình trạng lệch bội, phá vỡ rất nhiều trạng thái cân bằng trong các tế bào.)
Xác định giới tính ruồi giấm cũng khác với xác định giới tính của động vật có vú theo một số cách khác. Đầu tiên, xác định giới tính bắt đầu ngay khi thụ tinh, và không có giai đoạn “hai khả năng” (sẽ nói phần sau). Hơn nữa, hormone không chịu trách nhiệm cho các đặc điểm giới tính; thay vào đó, mỗi tế bào trong phôi cảm nhận tỷ lệ X: A, kích hoạt kiểu phiên mã đặc trưng cho cái hoặc đực. Các phân tích microarray chỉ ra rằng sự khác biệt đặc hiệu giới tính trong biểu hiện gen là khá rộng. Trên thực tế, khoảng 30% gen của ruồi giấm đã được phát hiện là có sự thiên lệch theo giới tính trong biểu hiện (Parisi et al., 2004).
Xác định giới tính ở chim
Ở chim, giới tính được xác định bởi nhiễm sắc thể được gọi là Z và W, và con cái là giới dị giao tử. Ban đầu, rõ ràng có sự khác biệt đáng chú ý trong các cơ chế được sử dụng để xác định giới tính ở chim và động vật có vú. Hãy nói về gà, các nhà khoa học không thể tìm thấy một bản sao nào của gen SRY cần thiết cho việc xác định tinh hoàn như ở động vật có vú, vì vậy họ đã tìm kiếm sự tương đồng của các gen khác cần thiết cho sự hình thành tinh hoàn của động vật có vú. Những nghiên cứu này đã dẫn đến việc phát hiện ra gen Dmrt1 trên nhiễm sắc thể Z của gà (Nanda et al., 1999).
Ở động vật có vú, sản phẩm gen Dmrt1 là thành viên quan trọng của con đường hình thành tinh hoàn do SRY khởi xướng. Hai bản sao của gen Dmrt1 là cần thiết cho sự phát triển tinh hoàn, ngay cả khi có mặt SRY. Ở động vật có vú, cả hai giới đều có hai bản sao Dmrt1, vì nó nằm trên một bộ nhiễm sắc thể (cụ thể là nhiễm sắc thể 9). Ngược lại, ở gà, chỉ có con đực sở hữu hai bản sao Dmrt1, vì nó nằm trên nhiễm sắc thể Z.
Yếu tố thực sự kích hoạt con đường hình thành tinh hoàn ở gà vẫn chưa được biết. Lớp Chim cũng khác biệt đáng kể với động vật có vú ở chỗ hai gen duy nhất trên nhiễm sắc thể W, FET1 và ASW, là cần thiết cho sự phát triển của con mái. Chức năng của FET1 chưa được biết, nhưng nó được thể hiện trong các tuyến sinh dục dẫn đến thời điểm phân biệt giới tính (Smith & Sinclair, 2004). Giống như động vật có vú, gà cũng có một tuyến sinh dục chưa xác định cho đến khoảng ngày thứ tư của quá trình phát triển, sau đó buồng trứng hoặc tinh hoàn bắt đầu hình thành. Hormone sau đó điều phối sự phát triển của các đặc điểm giới tính khác. Tuy nhiên, trái ngược với động vật có vú, estrogen được yêu cầu sớm hơn trong việc xác định giới tính ở gà và trên thực tế là cần thiết cho sự hình thành buồng trứng. Trên thực tế, gà trống có thể được chuyển đổi thành con mái nếu trứng được tiêm estrogen ở giai đoạn phát triển nhạy cảm.

Gà lưỡng tính, một nửa là trống, một nửa là mái. Ảnh: The Roslin Institute
Cơ chế xác định giới tính ZZ-ZW không giới hạn ở chim. Trong các loài động vật có xương sống, một hệ thống xác định giới tính tương tự đã được xác định ở các loài bò sát, cũng như ở một số loài cá và động vật lưỡng cư.
Xác định giới tính ở động vật có vú
Ở động vật có vú, sự hiện diện của nhiễm sắc thể Y quyết định giới tính. Thông thường, các tế bào từ con cái chứa hai nhiễm sắc thể X và các tế bào từ con đực chứa nhiễm sắc thể X và Y. Đôi khi, có những người được sinh ra với số lượng nhiễm sắc thể giới tính bất thường và giới tính của những người này luôn được xác định bởi việc có hay không có mặt nhiễm sắc thể Y. Do đó, những người có kiểu nhân 47, XXY và 47, XYY là nam, trong khi những người có kiểu nhân 45, X và 47, XXX là nữ. Con người có khả năng chịu đựng được số lượng nhiễm sắc thể giới tính quá mức do bất hoạt X và đồng thời nhiễm sắc thể Y của con người khá ít gen.
Nhiễm sắc thể Y và Sry
Mặc dù vai trò của nhiễm sắc thể Y trong xác định giới tính của động vật có vú đã được biết đến từ đầu thế kỷ XX, nhưng mãi đến năm 1959, các nhà khoa học mới có thể xác định vùng nhiễm sắc thể Y kiểm soát quá trình này (McLaren, 1991). Sau đó, nhà nghiên cứu David C. Page đã phân tích nhiễm sắc thể của những người đàn ông XX giới tính đảo ngược, những người hiếm hoi trông giống đàn ông nhưng có hai nhiễm sắc thể X thay vì một nhiễm sắc thể X và một nhiễm sắc thể Y. Sử dụng phương pháp lai DNA với các đầu dò tương ứng với các vùng khác nhau của nhiễm sắc thể Y, Page phát hiện ra rằng những người này mang gen thuộc về vùng 140 kilobase trên cánh ngắn của nhiễm sắc thể Y (Hình 1). Có lẽ, khu vực này đã được chuyển đến nhiễm sắc thể X do chuyển đoạn (Trang et al., 1985). Các thí nghiệm sau đó đã thu hẹp khu vực này (McLaren, 1991) và phát hiện ra rằng Sry là gen chính xác định giới tính. Do đó, sự hiện diện của vùng này trên nhiễm sắc thể Y là đủ để gây ra sự phát triển của nam giới (Koopman et al., 1991).
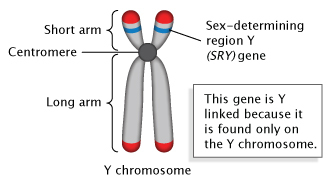
Vị trí của Sry trên nhiễm sắc thể Y của người. © 2014 Nature Education Adapted from Pierce, Benjamin. Genetics: A Conceptual Approach, 2nd ed.
Cơ chế phân tử xác định giới tính sơ cấp
Nhưng khoan hãy nhắc đến Sry vội, vì câu chuyện không hẳn bắt đầu ở đó. [Hãy lấy người làm ví dụ] Câu chuyện bắt đầu khi tuyến sinh dục của phôi vẫn còn ở giai đoạn “hai khả năng” (bipotential gonad), vẫn chưa có quyết định rằng phôi sẽ phát triển thành nam hay nữ, điều này tiếp diễn đến tận tuần thứ 6-7 của phôi. Các gen mã hóa nhân tố phiên mã như Wt1, Lhx9, GATA4, and Sf1 được biểu hiện và sự mất chức năng của bất kỳ một trong các gen này sẽ gây sự phát triển tuyến sinh dục bất thường. Sau đó lựa chọn mới được đưa ra:
+ Nếu không có nhiễm sắc thể Y, các nhân tố phiên mã và tín hiệu cận tiết này được cho là kích hoạt sự biểu hiện của Wnt4 và Rspo1. Lộ trình Wnt tạo ra nhân tố điều hòa phiên mã b-catenin. b-catenin thực hiện 3 chức năng bao gồm tác động tích cực trở lại Wnt4, b-catenin; khởi động lộ trình phát triển buồng trứng bằng và ức chế sự biểu hiện Sox9, một gen quan trọng cho sự phát triển tinh hoàn (Bernard et al. 2008; Lau and Li 2009).
+ Khi có nhiễm sắc thể Y, cũng chính các nhân tố này trong tuyến sinh dục “hai khả năng” kích hoạt gen Sry trên Y. SRY thúc đẩy sự biểu hiện của Sox9 (Bradford et al. 2009b; Sekido and Lovell-Badge 2009). Cả hai gen này cũng cùng ngăn chặn chiều hướng hình thành buồng trứng, có lẽ bằng việc chặn con đường b-catenin (Bernard et al. 2008; Lau and Li 2009).
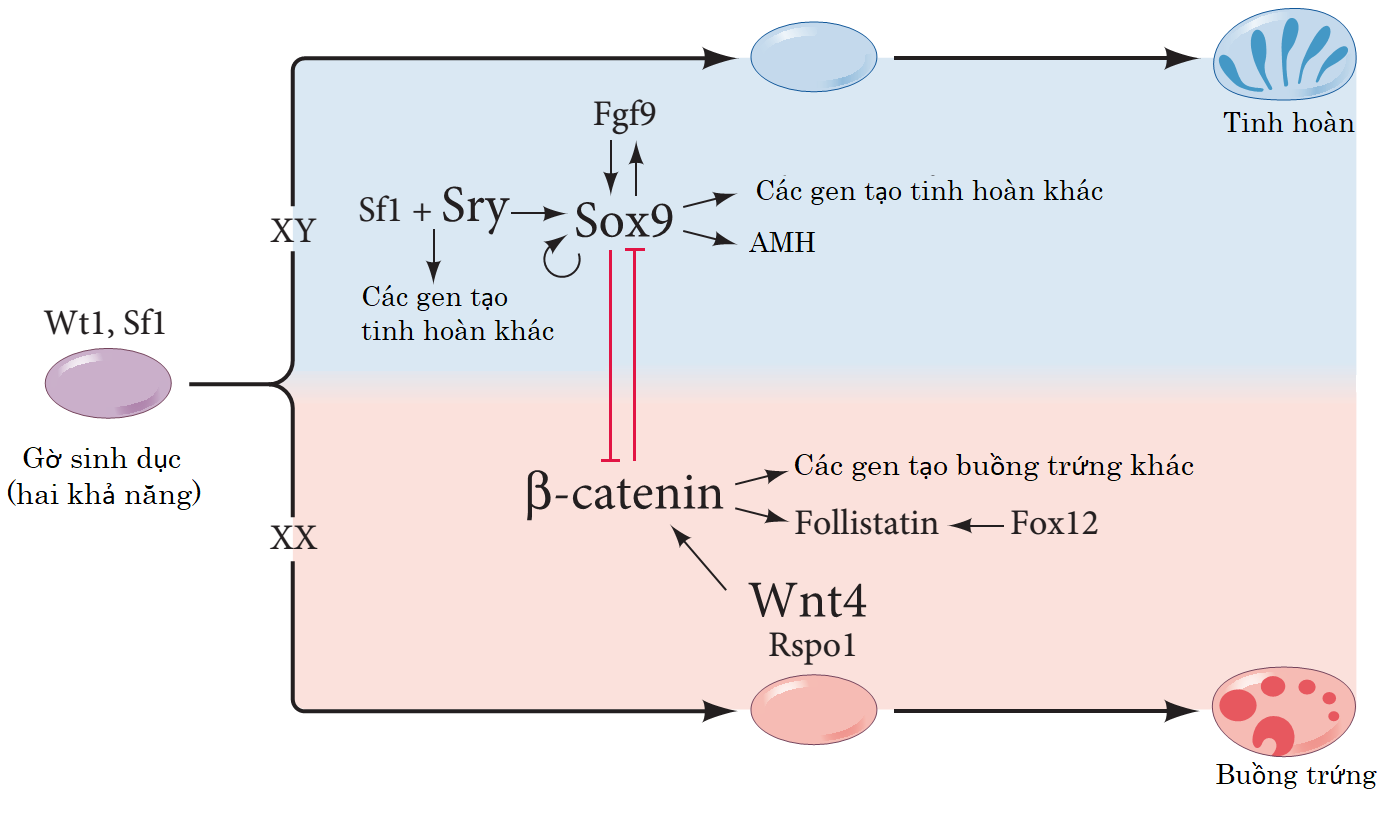
Những gen tham gia vào việc quyết định giới tính trong thời kỳ tuyến phôi. (Sekido and Lovell-Badge
2009; McClelland et al. 2012)
Trong giai đoạn phôi, gen Sry kích hoạt con đường hình thành tinh hoàn vào khoảng tuần thứ bảy của sự phát triển. Trước thời điểm này, tuyến sinh dục phôi thai “không phân hóa” hay “hai khả năng”, nghĩa là nó có khả năng phát triển thành tinh hoàn hoặc buồng trứng. Tương tự như vậy, phôi sớm có hai hệ thống ống dẫn, ống Wolffian và Müllerian, tương ứng có khả năng phát triển thành vùng sinh sản nam và nữ. Một khi sản phẩm gen SRY kích thích tuyến sinh dục phát triển thành tinh hoàn, tinh hoàn bắt đầu sản xuất hai loại hormone, testosterone và hormone chống Müllerian (AMH). Testosterone và một trong những dẫn xuất của nó, dihydrotestosterone, gây ra sự hình thành các cơ quan khác trong hệ thống sinh sản nam, trong khi AMH gây ra sự thoái hóa của ống Müllerian. Ở nữ giới, không chứa protein SRY, con đường hình thành buồng trứng được kích hoạt bởi một bộ protein khác. Buồng trứng phát triển đầy đủ sau đó tạo ra estrogen, kích hoạt sự phát triển của tử cung, ống dẫn trứng và cổ tử cung từ ống Müllerian.
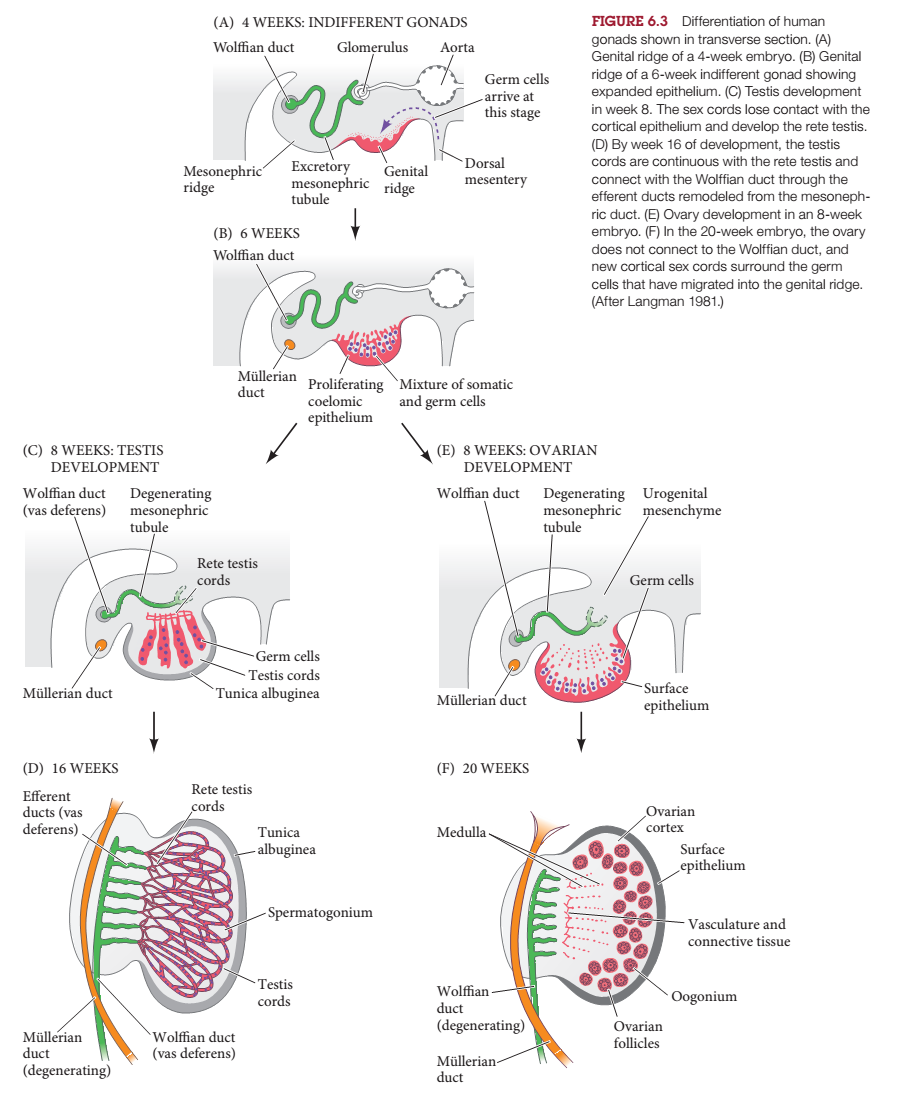
Sơ đồ phát triển tuyến sinh dục nam và nữ trong thời kỳ phôi thai.
Con đường tạo tinh hoàn: Sry và Sox9
Ở người, gene Sry nằm ở đầu mút phía vai ngắn của nhiễm sắc thể Y. Protein SRY gổm 223 axit amin, chứa một vùng bám DNA gọi là HMG box (high-mobility group), là một nhân tố điều hòa phiên mã, quyết định sự hình thành tinh hoàn. Những ghi nhận thực tế đã giúp chứng minh vai trò của Sry:
+ Người có cánh ngắn của Y mà không có cánh dài vẫn là nam nhưng có cánh dài mà không có cánh ngắn thì là nữ.
+ Sry thấy ở nam XY bình thường và nam XX hiếm
+ Sry không có ở nữ XX bình thường hay nữ XY.
Còn một gen thực ra cũng rất quan trọng đối với sự hình thành giới tính nam. Gene Sox9 (SRY-Box 9) nằm trên vai dài nhiễm sắc thể 17, mã hóa một nhân tố phiên mã giả định cũng chứa HMG box. Protein SOX9 biểu hiện ở mầm tuyến sinh dục đực hơi muộn hơn SRY. Trong khi gen xác định giới tính Sry đặc trưng ở thú, Sox9 có tất cả ở động vật có xương sống. Nhiều thí nghiệm đã chứng minh vai trò của gen này trong việc hình thành giới tính, thậm chí bất kể là có SRY hay không, ví dụ:
+ Người có XX khi thêm một bản sao Sox9 thì người đó là nam, kể cả là không có Sry (Huang et al. 1999; Qin and Bishop 2005); chuột XX chuyển gene Sox9 cũng phát triển tinh hoàn.
+ Loại bỏ gen Sox9 ở tuyến sinh dục chuột có cặp nhiễm sắc thể XY, chuột phát triển thành chuột cái, kể cả là có Sry trên Y (Barrionuevo et al. 2006).
Với những dữ kiện như vậy, có cơ sở để cho rằng Sox9 dường như là gene xác định giới tính chủ chốt và còn xuất hiện sớm hơn Sry trong tiến hóa (Pask and Graves 1999). Biểu hiện Sox9 đặc biệt tăng khi có sự kết hợp của Sry và Sf1 trong các tế bào Sertoli tiền thân. Nói cách khác, Sry có lẽ chỉ đơn thuần là kích hoạt Sox9 trong một thời gian rất ngắn lúc đầu.
Sự hình thành buồng trứng: Wnt4 và R-spondin1
Sự hình thành giới tính nữ cũng không chỉ đơn thuần dựa vào câu hỏi có Sry hay không. DAX1 là một gene nằm trên vai ngắn của nhiễm sắc thể X quyết định sự hình thành buồng trứng. Dax1 cũng biểu hiện trong tuyến sinh dục sơ khai cùng thời gian với Sry, làm ức chế sự biểu hiện của Sf1. Cá thể có XY mà mang 2 bản sao của Dax1 trên X thì vẫn phát triển thành cái, kể cả có Sry.
Wnt4 trên nhiễm sắc thể 1 biểu hiện ở tuyến sinh dục sơ khai của cả 2 giới; nhưng sau đó chỉ biểu hiện được ở tuyến sinh dục đang hình thành buồng trứng. Chuột XX chuyển gene mất Wnt4 không tạo được buồng trứng mà biểu hiện Sox9, enzymes tạo testosteron và AMF (Vainio et al. 1999; Heikkilä et al. 2005). Trong khi đó Rspo1 hoạt động cùng Wnt4 để tạo β – catenin – giúp phát triển buồng trứng và ức chế biểu hiện Sox9. Ở người mang XX mà có đột biến RSPO1 trở vẫn phát triển thành nam; ngược lại, người XY tồn tại lặp đoạn chứa cả Wnt4 và Rspo1 trên nhiễm sắc thể 1 thì vẫn sẽ là nữ.
Nói tóm lại, việc xác định và phát triển giới tính ở thú nói chung và ở người nói riêng không chỉ phụ thuộc vào nhiễm sắc thể giới tính, mà còn phụ thuộc nhiều vào một số gen trên nhiễm sắc thể thường.
Các khác thường về giới tính ở người
Cũng chính vì những cơ chế rắc rối kể trên mà ở người cũng hình thành những giới tính sinh học đặc biệt, chúng ta sẽ thật bối rối khi nhắc tới khái niệm “nam” hay “nữ”. Ví dụ, trường hợp lưỡng tính giả nữ, về mặt di truyền là nữ (XX) nhưng biểu hiện là nam; lưỡng tính giả nam: về mặt di truyền là nam nhưng biểu hiện là nữ.
Một kiểu giới tính khác thường nữa là lưỡng tính thật, họ là thể khảm 46/46 XY/XX trong 1 tuyến sinh dục, tạo ra cả cấu trúc buồng trứng tinh hoàn và cả 2 loại hormone được tạo ra. Một khả năng khác cũng có thể xảy ra, khi một đôi tuyến sinh dục thì một bên là 46 XX, một bên tuyến là 46 XY, như vậy giống con gà lưỡng tính, một nửa nam một nửa nữ. Thể khảm phát triển từ giai đoạn phôi, còn trưởng thành mà cấy ghép tế bào để tạo thể khảm thì cũng không tạo ra lưỡng tính thật. Người chuyển giới sử dụng hormone sinh dục nam/nữ để tạo ra kiểu hình thứ cấp tương ứng.
Đứa bé sinh ra là gái nhưng lớn lên thành chàng trai? Bản chất là nam có tuyến sinh dục nam điển hình nhưng hormone testosteron chưa đủ hoặc bất thường receptor nên ban đầu tưởng là “gái”. Khi dậy thì testosteron biểu hiện mạnh đủ để thay đổi kiểu hình bên ngoài trở về đúng là nam. Vậy lúc đầu nữ chỉ là giả.
Xác định giới tính và môi trường
Như đã đề cập trước đó về Talaeporia tubulosa, các yếu tố môi trường đôi khi có thể đóng một vai trò quan trọng trong việc xác định giới tính. Côn trùng không phải là trường hợp đặc biệt – giữa các loài động vật có xương sống, nhiệt độ cũng có ảnh hưởng mạnh mẽ đến việc xác định giới tính ở một số nhóm bò sát nhất định. Ví dụ, trong các loài bò sát, cá sấu và hầu hết các loài rùa, giới tính được xác định bởi nhiệt độ ấp trứng. Có một số biến thể về chủ đề này. Ở loài rùa cá sấu, Macroclemys temminckii, ấp trứng dưới 22 ° C hoặc trên 28 ° C sẽ sinh ra con cái, trong khi ủ ở nhiệt độ trung gian tạo ra con đực là chủ yếu. Cá sấu Mỹ cho thấy một sự phụ thuộc hai pha tương tự vào nhiệt độ, nhưng đường cong được chuyển sang nhiệt độ cao hơn. Ở rùa ao châu Âu, Trạchemys scripta, nhiệt độ ấp trên 30°C sinh ra tất cả con cái, trong khi nhiệt độ ấp dưới 25 ° C tạo ra tất cả con đực. Ở 28,5 ° C, số lượng đực cái bằng nhau được sinh ra.
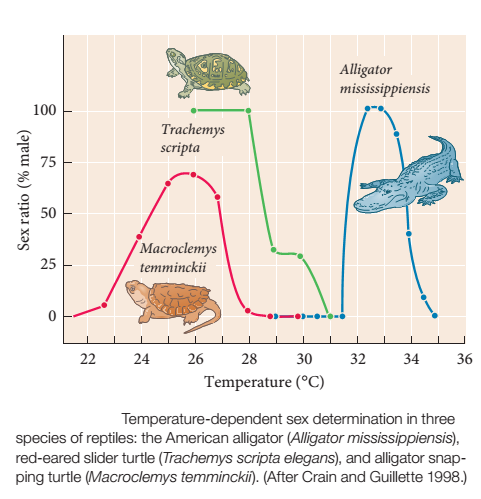
Sự phụ thuộc của giới tính con non bởi nhiệt độ ấp trứng. (Crain and Guillette 1998.)
Thành phần phụ thuộc nhiệt độ của con đường xác định giới tính đã được nghiên cứu rất chi tiết ở loài rùa châu Âu, Emys orbicularis. Ở E. orbicularis, mấu chốt dường như là sự tổng hợp enzyme aromatase, giúp chuyển đổi androgen, như testosterone, thành estrogen. Ở nhiệt độ cao hơn, hoạt động aromatase tăng lên tạo ra nhiều estrogen hơn, điều này làm lệch tỷ lệ giới tính về phía tạo con cái.
Khi so sánh các cơ chế khác nhau để xác định giới tính giữa các loài, rõ ràng sự tiến hóa đã tạo ra nhiều cách thức để tạo ra các giới tính khác nhau. Sinh sản hữu tính có giá trị thích nghi rất lớn đối với một loài, bởi vì nó tạo nên sự biến đổi di truyền mới cho quần thể trong mỗi thế hệ mới. Nhiễm sắc thể đóng vai trò quyết định ở hầu hết các loài, nhưng ngay cả như vậy, các yếu tố môi trường cũng để lại những gợn sóng nhất định bổ sung vào quá trình phát triển.
iceberg (biên tập)
tapchisinhhoc.com
